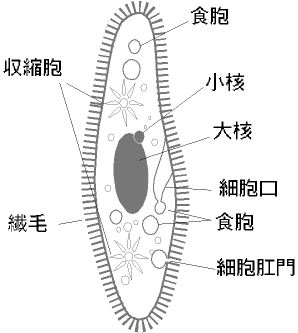
Key to the species
細胞は円筒形ないし葉巻形; 先端は丸く, 後端は尖る; 先端から細胞長の2/3の部分が最も幅広い
小核は1個で大きい (compact type, 8 μm); 収縮胞は2個; 長さは 170-290 μm .......................
P. caudatum
小核は2個かそれ以上 (vesicular type, ~2 μm)
後端部は caudatumほどは尖らない; 小核は2個; 収縮胞は2個; 長さは 80-170 μm ..............
P. aurelia
後端部は caudatumと同様に尖る; 小核は3-4 個 (up to 7); 収縮胞は2-3個 (最大 6個);
長さは180-310 μm (最大 400 μm) .............................................................
P. multimicronucleatum
細胞は短く幅広い, 背腹方向にややつぶれている; 後端部も丸い
細胞内にクロレラを共生させている; 先端部は平坦, 後端部は丸い;
小核は1個で大きい (compact type, ~7 μm); 長さは 85-150 μm .............................................
P. bursaria
共生クロレラはいない
先端はやや丸い, 後端も丸い; 小核は通常1個 (まれに最大3個, compact type, 4-7 μm);
収縮胞は2個; 長さは 70-90 μm ...................................................................................
P. trichium
海水性のものもいてそれらの収縮胞は不明瞭, radial canal のみが目立つ
細胞の中央部が最も幅広い; 他と異なり遊泳時に右方向に回転する; 収縮胞は2個;
小核は通常2個 (まれに最大5個, vesicular type, 3-5 μm); 長さは 110-140 μm ..........
P. calkinsi
細胞口は細胞体のほぼ中央にある; 収縮胞は2個;
小核は通常4 (1-8)個 (vesicular type, 3-4 μm); 長さは 70-110 μm .........................
P. polycaryum
細胞口は細胞体の中央より後端部側にある; 収縮胞は2個;
小核は通常3-4 (-8)個 (vesicular type, 4-5 μm); 長さは 120-210 μm .........................
P. woodruffi
(From Wichterman, 1986)
|
 1
1
 2
2
 3
3
 4
4
 5
5
 6
6
 7
7
 8
8
 9
9
 10
10