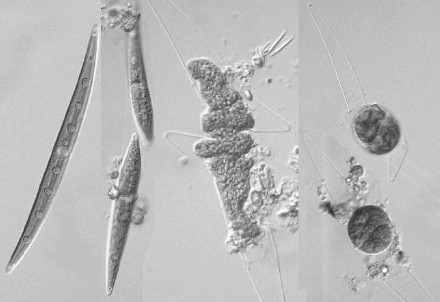
繊毛虫は遊泳しながら分裂するため,分裂中の細胞にであっても連続して分裂の様子を観察するのは難しい。しかし,分裂中の細胞は他の細胞に比べて遊泳が活発ではないので,試験管培養をすると試験管の底に分裂中の細胞がたくさん集まるようになる。そのため,そこを狙えば分裂中の細胞を探しやすい。
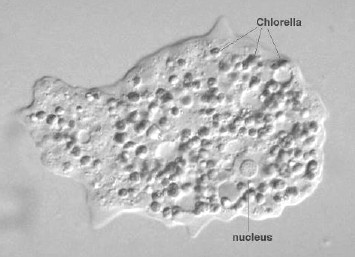
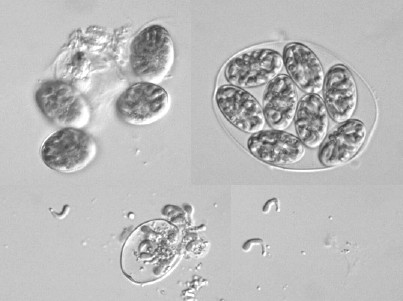
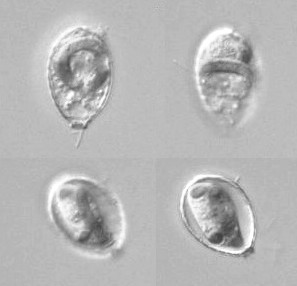
繊毛虫は細胞表層に細胞口や複雑な形の構造があるが,分裂の際にこれらがどのようにして「複製」されるかに興味がもたれ,長年研究されている。また,繊毛虫の中には,特殊な分裂をするものがいる。
ツリガネムシ
類は分裂の際,片方の細胞が柄を離れて泳ぎだす。吸管虫と呼ばれるグループは野外では比較的希だが,分裂は「出芽」と呼ばれる特種な形式をとる。
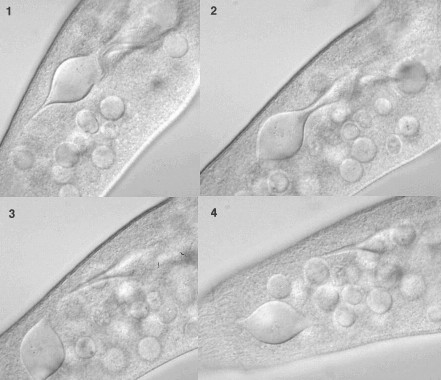
Tokophrya
などでは分裂後の片方の細胞がもとからあった細胞の中にできる(内生芽という)。分裂終了後,この内部にできた細胞はもとからあった細胞にあいた孔から出て繊毛を使って遊泳する。しばらく遊泳すると固形物に付着するようになるが,その後,繊毛が消失し,吸管虫特有の形態に変化する。
|

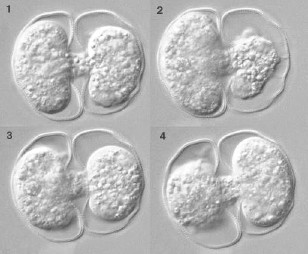
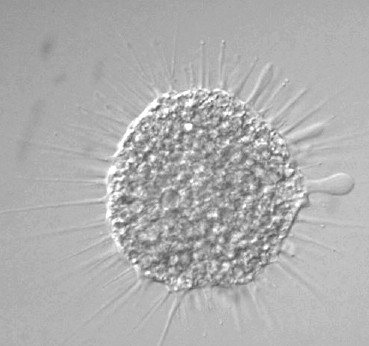
アワセオオギ(Micrasterias crux-melitensis )の分裂
分裂は「半保存的」に起こる。中央のくびれ部分から分裂が始まり,それぞれの分裂面から新しい半細胞(くびれ部分で分けた細胞の半分)が形成される。新しくできつつある半細胞は当初薄い膜で囲まれている。この中で,半細胞が成長して片側にあるもとからあったものと同じ形の半細胞ができる。2つの娘細胞は,互いの半細胞が完成するまで接着していることが多い。
|