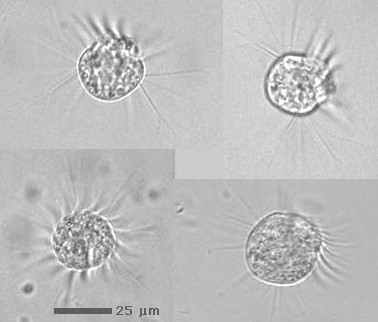
停止した状態と瞬間的な高速移動をくり返す。停止した時を狙って撮影した。
| 原生生物の採集と観察 |
| もどる | 目 次 | すすむ |
2 原生生物の観察法
2)生物ごとの観察法
|
原生生物の多くは,生きた状態でないとその特徴がつかめないものが多い。そのため,原生生物を観察する際には,生細胞をいかに自然なまま観察・記録するかが重要なポイントとなる(勿論,細胞分裂の際の核変化や細胞表面の微細構造を見るためには,細胞を固定・染色したり,内部の原形質を除去する等の処理が必要になる)。
ここでは,各生物群ごとに観察をする際のコツ,ないしは留意点について紹介する。 |
| 繊毛虫 |
| 繊毛虫の多くは,動き(遊泳行動)が活発なため,顕微鏡で観察するには,いかにしてその遊泳を止めるか,ないしは,遊泳速度を遅くするかが問題になる。細胞遊泳を止める方法としては,例えば ゾウリムシ であれば,細胞を粘稠な液体中に入れたり,細胞を「麻酔」するなどがある(詳しくは4章を参照)。ただし,このような方法で遊泳を止めると,種類によっては,傷ついたり,種の特徴となっている外形が崩れてしまうものもいる(例:ハルテリア等)。一番簡単なのは,カバーグラスの周囲から水分が蒸発することで,カバーガラスとスライドガラスの隙間が徐々に狭まっていくのを利用する方法である。これにより細胞体の大きなものから順番に2枚のガラス板の間に挟まって動けなくなる時期が訪れるので,その時を狙って観察するのである。もちろん,その後,さらに時間がたつとガラスの隙間がさらに狭まるため,細胞は変形しやがては破壊されてしまうが,細胞体があまり変形しない段階までならば,比較的自然なままの細胞の様子を詳しく観察することができる。 |
停止した状態と瞬間的な高速移動をくり返す。停止した時を狙って撮影した。 |
|
また,細胞が2枚のガラス板に押されて扁平になると,それまで見えにくかった細胞内部の核や収縮胞,食胞などの微細構造が観察しやすくなる。核は通常は染色をしないと見えないように思われがちだが,細胞を平たくすると生きたままでも容易に観察することができる。
動きが活発とはいえ,繊毛虫の多くは常に動き回っているわけではない。カバーガラスをかぶせた直後などはその刺激により一時的に活発に動き回るが,やがて落ち着いてきて動きが緩慢になるので,あわてずに時間をおいてから観察するのがよい。中には ゾウリムシ のように繊毛を介して固形物に付着して遊泳を停止するものもいる。このような状態の細胞では,後述するように,細胞口から餌のバクテリアを取り込んで食胞を形成する様子や,その食胞が細胞内を動き回る様子(原形質流動),細胞口門から未消化物を排泄する様子,あるいは収縮胞の動きなどを観察することができる。 中には,いつまで待っても遊泳速度が衰えないものや, スピロストマム のように,絶えず変形して正確な外形をとらえきれないものもいる。このような場合は,可能であれば,顕微鏡用のビデオカメラで動画を記録し,後で画面を止めて観察したり,静止画として保存するのがよい。 |
| 肉質虫 |
| 肉質虫は大きく有軸仮足虫類(太陽虫の仲間)と根足虫類(アメーバの仲間)の2グループからなる。いずれも動きは繊毛虫ほど活発ではないので,特別な処理をせずともそのまま顕微鏡で観察できる。とくに有軸仮足虫は動きが緩慢なので観察しやすい。ただし,長い軸足を放射状に出しているため,カバーグラスをかぶせて,余分な水分を除こうとすると軸足が折れ曲がってしまい細胞体も変形してしまうことが多い。そのため,有軸仮足虫を観察する際は,スライドガラスとカバーガラスの間に通常より多めに水を残しておく必要がある。ただし,こうすると他の原生生物にはピントが合いにくくなる。 |
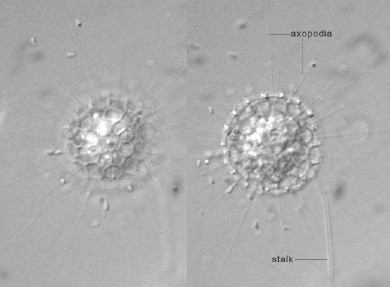 殻を持った太陽虫の一種。比較的希。殻の内部に細胞体があり,そこから殻の隙間を通って軸足が伸びる。細胞分裂は殻の中で起こる。 |
| 一方,根足虫類はやや動きが活発だが,それでも顕微鏡下(x 100〜 x 400)でゆっくり観察できる程度の速さでしかない。根足虫類は,その移動時の細胞の形(仮足の形)が種を区別する重要な判定基準となる。たとえば,筒状( Amoeba とその仲間),平らな三角形( Mayorella 類),糸状( Gromina 類)などである。そのため,太陽虫同様,観察の際には,カバーグラスを細胞体に押し付けないよう注意しなければならない。また,すべてではないが,中には光に敏感なものもいる。それらは写真撮影などのために光量を多くすると細胞が刺激されて動きが止まったり,原形質流動の方向転換が頻繁に起きるため,アメーバ固有の移動形が崩れてしまう。そこで,観察する時は光量を落とし,撮影の瞬間だけ光量を上げるようにする。 |
| 多くの根足虫類は,スライドグラスに移した段階では細胞体が凝縮しているが,しばらく放置すると新しい環境に慣れて移動するようになる。しかし,中にはかなりの時間をおかないと細胞体が正常な形に回復しないものもいる。そのような場合は,水分が蒸発して乾燥しないように,カバーグラスの周囲を何かで囲う必要がある。著者は,カバーグラスをかぶせる前に,スライドグラスに接する側の周辺部に毒性のないグリースを塗っておく方法を採用している。その際,塗るグリースの厚みを変えることで,観察しようとする細胞体の大きさに合わせて2枚のガラス板の間隔を調整することもできる。この方法だと,一日以上の長期の観察が可能である。これにより,1日目には観察できなかった長い仮足を持つアメーバ(顆粒状根足虫類の仲間)を翌日になって観察できたことがある。 |
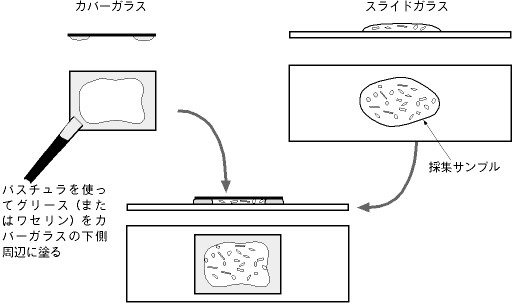
採集サンプルを長期間観察する方法 |
| 鞭毛虫 |
| 鞭毛虫類の多くは非常に小型のため,双眼実体顕微鏡ではその存在すら気がつかないものが多い。その中で比較的細胞が大きく観察しやすいのが, ミドリムシ 類と渦鞭毛虫(渦鞭毛藻ともいう)類である。 |
| 渦鞭毛虫類は淡水では希なグループだが,ときおり大量発生していることもある。ほとんどは硬い殻(鎧板,theca)を持っているタイプで,この殻の形で種の同定が行なわれる。鎧板の形は,細胞が死んで殻だけが残っているものがあれば観察しやすい。生きているものはさかんに遊泳するし,細胞内にたくさんの葉緑体やその他の顆粒を保持しているので,そのままでは観察しずらい。生きているものを観察するには,繊毛虫の場合と同様,遊泳を遅くする処置をするか,カバーグラスで軽く抑えて動きを止める必要がある。ただし,殻が壊れやすいので押し付ける場合は注意が必要である。 |
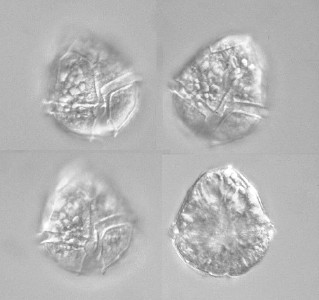 渦鞭毛虫ペリディニウム (Peridinium bipes) |
|
ミドリムシ
類は,小型のものは動き(遊泳行動やミドリムシに特有のユーグレナ運動)が活発な種類が多い。大形の種類はまれに遊泳することもあるが通常は水底でじっとしているので,細胞内部の観察材料としては,野外で見つけやすい大型種が向いている。
ミドリムシの細胞表面には螺旋状の縞模様がある(大型種ではハッキリ見える)。細胞内には,核や葉緑体の他に棒状やリング状をしたパラミロン粒が数個あるいは小さく身短いものが多数見られる。葉緑体は,円盤状のものが細胞表層部にあるタイプと,リボン状ないしは短冊状のものが細胞中心部から周辺部に向って広がっているタイプのものがある。後者は細胞内の他の顆粒がじゃまで見えにくいが,円盤状のものは細胞表層にピントを合わせることで比較的容易に観察できる。 |
| ミドリムシは,走光性を観察するのによく用いられるが,走光性を見るためには,遊泳活動が活発な細胞が大量に必要となる。したがって,大量培養が難しい大型の種類は不向きで,やはり Euglena gracilis のような大量培養が容易で,かつ遊泳活動が活発なものが適している。とはいえ大量培養するとなると,それなりに手間がかかる。しかし,既述したように,ミドリムシは野外で大量に(かつ高密度で)増殖している場合が多い。それらは比較的小型種で遊泳活動が活発なものが多いので,これらも走光性の実験に利用できるはずである。事前に大量増殖している場所を何ケ所か確認しておけば,実験前にそれらの「野外にある培養器」から必要量を採取して利用できるだろう。 |
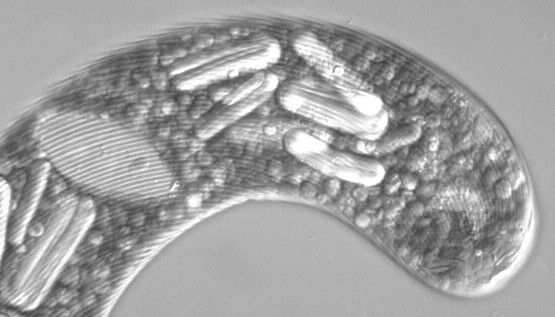 大型のミドリムシ(Euglena gigas) 細胞表層にある螺旋状の縞模様が見える。中央部に透明な楕円形の核が,先端部に大きな眼点が見える。その間に棒状(実際はリング状のものが潰れて棒状に見えるだけ)のパラミロン粒が何本か,さらに顆粒状の粒子も多数見える。葉緑体は縞模様のある表皮(ペリクル)の直下に小さな円盤状のものが多数ある。ただし,この画像は白黒のため葉緑体は見分けにくい。核の上に1つだけあるのが見える。 |
| 不等毛類 |
| 聞き慣れない言葉かも知れないが,不等毛類とは,従来,黄金色藻類( オクロモナス, ヒカリモ, シヌラ, マルロモナス 等),黄緑色藻類( フシナシミドロ, トリボネマ 等),ラフィド藻類( ゴニオストマム 等),珪藻類と呼ばれてきた生物群をひとつにまとめた呼び名である。その名の通り,細胞の先端部から長い鞭毛と短い鞭毛が伸びている時期が生活環の一部にみられるのが特徴とされる。各グループには葉緑体を持つものと持たない生物が混在しているが,葉緑体を持つものは,遠い祖先が他の藻類を「 細胞内共生 」させたものであることが知られている。 |
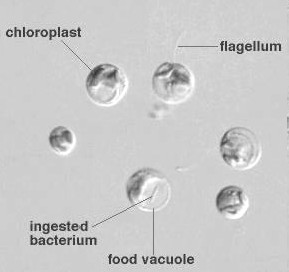 鞭毛を持ち動き回り,鞭毛基部にある細胞口でバクテリアを捕食する。しかし,同時に葉緑体もあり,光のみでも増殖が可能。無菌培養もできて大量培養が容易。ただし,細胞は小さい。 |
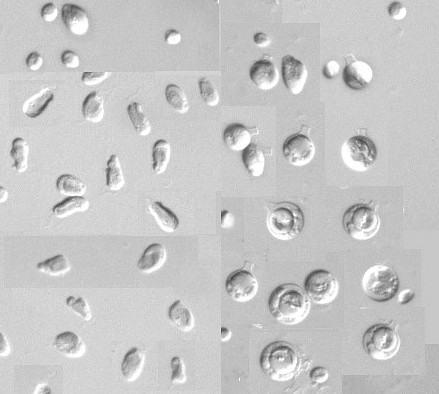 左側が増殖期の細胞,内部に黄色味がかった葉緑体が見える。鞭毛は1本のみ。右側はトックリ状のシスト。 |
|
黄金色藻類,黄緑色藻類,ラフィド藻類の3グループは比較的希な種類が多く,また,細胞体も小さいので観察には不向きである。文献でも上記の代表的な種類以外はほとんど紹介されていないが,小型のものはいたるところに生息している。オクロモナスや
ヒカリモ
は採集当初はほとんど見つからないが,シャーレなどにサンプルを入れ光を当てて放置しておくと,いつのまにか水の表面に多数増殖していることがある。
ヒカリモ
はもっとも小さい部類に属するが,これが水の表面をおおうと,光の反射が通常と異なるためすぐに気付くことができる。
一方,珪藻類は数も種類も多い。珪藻類は,細胞体を溶解して,殻の細部をみて分類を行なうため,文献には殻の形状のみが掲載されることが多い。しかし,既述したように(1章),生きた珪藻は滑走運動によりさかんに動き回っている。平面を比較的ゆっくりとした速度で移動するので,光学顕微鏡で観察するのに向いている。 |
| 緑藻類 |
| 緑藻類には鞭毛を持って泳ぐタイプ( クラミドモナスや ボルボックス の仲間)と,水底でじっとして動かないタイプ(クロレラ類,接合藻,他)がある。前者の細胞は一般に小さいので細胞内部の観察には向いていない。しかし, ミドリムシ と同様に,走光性の実験などには適している。また,単細胞性の クラミドモナスとその仲間は接合の観察に,群体で生活する ボルボックス の仲間は多細胞化の過程を見る材料として紹介されることが多い。ただし,接合を観察するには ゾウリムシ の場合と同様,それなりの準備がいる。群体で生活する種類は細胞分裂後に新しい群体が形成されるが,その過程では群体の反転現象(colony inversion)が起こるなど興味深い変化が見られる。 |
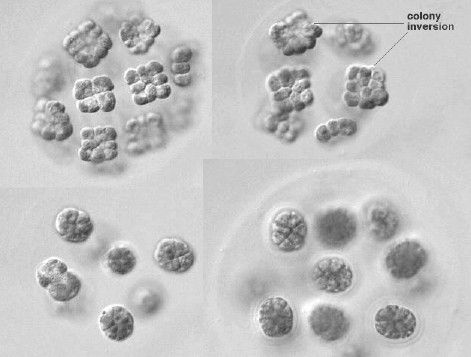 親群体を構成するすべての細胞が分裂に参加する。新しくできた娘群体は途中で「反転」を起こす。 |
| 一方,泳がないタイプの代表であるクロレラ類は細胞は小型のものが多い上,浮遊生活をするだけなので,細胞分裂過程以外にとくに観察すべきものはない。また,分裂は主に夜間に起こるので,観察しずらい。群体を形成する クンショウモ 等では,分裂の際に著しい形態変化が見られるが,細胞が小さいので観察しずらい。接合藻類は,一般に細胞体が大きいので,これも後述するように,原形質流動や細胞分裂の様子を観察するのに向いているが,接合を観察するには多少幸運に恵まれる必要がある。接合に遭遇できれば興味深い変化を観察することができる。 |
| もどる | 目 次 | すすむ |
| 原生生物の採集と観察 |