1 分子・形態レベルにみられる変異の飽和


| 第1回日本進化原生生物学研究会: 原生生物における種の実在性 | by 月井雄二 |
1 分子・形態レベルにみられる変異の飽和 |
|
1-1. 接合型グループ(旧称:シンジェン)は多系統か?
ゾウリムシの接合型グループは生物学的種ではない
原生生物ゾウリムシ(Paramecium caudatum)では,従来,接合型の特異性が異なるグループは互いに生殖的に隔離されている(はず)なので,それぞれは別種である,と考えられてきた。
しかし,私は接合型の遺伝解析を目的として接合型グループ間の雑種を作成したところ,その雑種に妊性がある(子孫を残す能力がある)ことを発見した(Tsukii & Hiwatashi 1983; Tsukii 1988)。これにより,グループ間では生殖的隔離が起きていない可能性が示唆された。
図1 ゾウリムシの接合と接合型グループ
相補的な接合型(EとO)を混ぜると繊毛を介して細胞凝集が起こり,やがて接合対ができる(左図)。 この接合型の組み合わせは同じ形態種内に複数存在する(右図)。
ミトコンドリアDNA,および,大核DNAのRAPD系統樹と接合型グループの分布は一致しない
その後,ゾウリムシから抽出したミトコンドリアDNA,および核DNAを使った分子系統樹を作成したところ,それらと接合型の分布はまったく一致しなかった(Tsukii 1994; Tsukii 1996)。
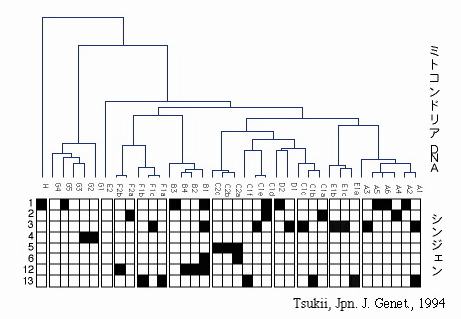
図2a ミトコンドリアDNAのRFLP系統樹と接合型グループ
図の上段はミトコンドリアDNAの制限酵素断片の比較を元に作成した系統樹でA1, A2などの記号は制限酵素断片位置の違いに基づくミトコンドリアDNAの種類を示している;下段は各ミトコンドリアDNAを持っていたゾウリムシが所属していた接合型のグループ(左脇に番号で表示;該当した場合を黒色で表している)。
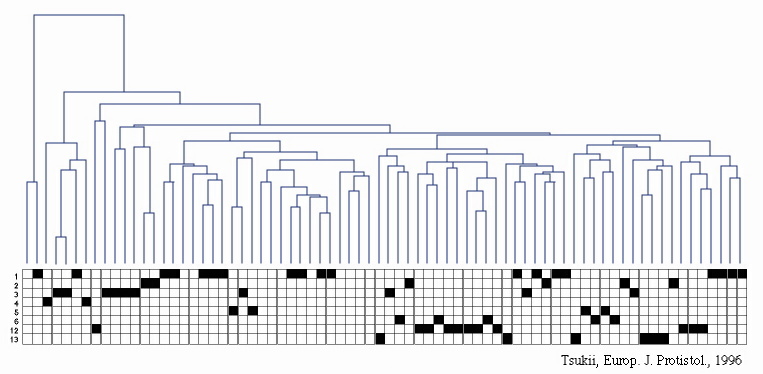
図2b 大核DNAのRAPD系統樹と接合型グループ
図の上段は,ゾウリムシから抽出した大核DNAを鋳型として作成したRAPD (Random Amplified Polymorphic DNA)系統樹;下段は各枝部分に該当したゾウリムシが所属していた接合型グループ(左脇に番号で表示)。
これらの結果は,各接合型グループは,いわゆる単系統(単一の祖先のみ由来する生物集団)ではなく多系統(系統的に異なる複数の祖先からなる集団)であることを強く示唆している。すなわち,ゾウリムシでは,祖先が異なるにもかかわらず,見かけ上,同じ接合型特異性を持つものが多数存在する(すなわち接合型に関して収斂進化が起きている)可能性が高い。
接合型の変異の中に他の接合型特異性を持つようになったものがいる
実際すでに,野外からそれと思われる接合型の変異(多型)もいくつか発見されている(Tsukii 1988; 堀ら 1988)。
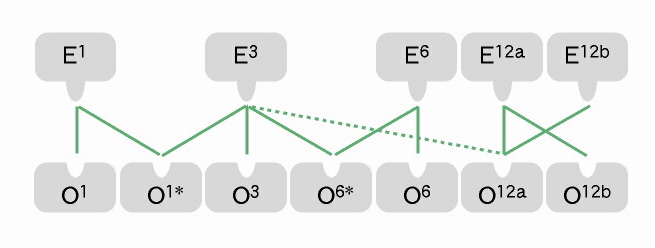
図3 接合型の多型
従来,接合型はEとOが1対1の関係にあると思われていたが, 詳しく調べると,中にはO1*のように2つのEタイプ(E1 & E3)と反応するものが発見された(月井)。 遺伝解析の結果,このO1*はO1の変異であることが判明した。
少なくとも2億4000万年前からゾウリムシはいた!
一方,近年の化石研究(Schöborn et al. 1999)から少なくとも2億4000万年前にはすでに今と形態的には変わらないゾウリムシが生息していたことが知られている。
ゾウリムシの接合型変異はすでに飽和している!?
P. caudatumでは形態的には単一種とされるが,既述した接合型の違いにより16の接合型グループに分かれている。しかし,P. caudatumが2億4000万年前から存在していたとすれば,その間に突然変異により分化した接合型グループの数がわずか16というのは数が少なすぎる。
考えられる理由は,接合型に関与する分子(接合型物質と呼ばれるタンパク質)の細胞接着に関与する接合型活性部位の構造が比較的単純で,その取りうる立体配置が16種類しか存在しえなかったのではないか,ということである(注1)。そのため,16のグループが誕生した後に起きた接合型の変異は,すべて既存のいずれかの接合型に収斂してしまい16以上には数が増えなかったのであろう。

図4 ゾウリムシの系統進化と接合型変異の飽和
ゾウリムシでは,接合型の数がすでに飽和状態にあり,その結果,各接合型への収斂進化が何度も起きている可能性がある。注1:接合型物質そのものはある程度のアミノ酸配列を持っているはずだが,接合型活性に関与する部位はそのごく一部のはずである。無論,一部とはいえ,数十個程度のアミノ酸配列は関与しているだろうから,それらが変異しうる可能性は10や20ではないだろう。しかし,この物質には,EタイプとOタイプという相補的な組み合わせが成立して始めてその機能が保持される,という制限条件が付いている。このため,E,Oいずれのタンパク質においてもその接合型活性部位のアミノ酸配列の変異が即,接合型特異性の変異として検出できるわけではない。結局は,接着する相手側のタイプが存在しなければならないので,接合型特異性の変異として検出されるものは限られてしまう。
もしかすると,同じ接合型(例えば,E1)と判定されるものでも,実際にはその接合型活性部位のアミノ酸配列には変異があるのかも知れないが(E1a, E1b, E1c),それが検出されるためには,相手側の接合型(この場合は,O1)にも,その違いを検出できる変異(O1a, O1b, O1c)が存在しなければならない。以前は,単純に既存の接合型(O1, O2, O3など)と反応するかどうかだけで調べていたため,このような同じ接合型の変異についてはわからなかったのである。
しかし,E, Oそれぞれ同じ接合型(例えば,E1とO1)とされる野外株を複数揃え,可能なすべての組み合わせで混ぜ合わせると,実際に,図3のような接合型の多型現象が発見されたのである。このことから,接合型の変異は,接合型グループの数(=互いに排他的関係にあるE, O接合型活性部位の組み合わせ)以上に多く存在することが示唆される。
| Copyright 2004 by Yuuji Tsukii |
|
