月井 雄二(法政大学 自然科学センター)
第1回日本進化原生生物学研究会,金沢大学理学部, 2003年6月28-29日
URL, http://square.umin.ac.jp/jsep/
 |
タイトル:原生生物における種の実在性について |
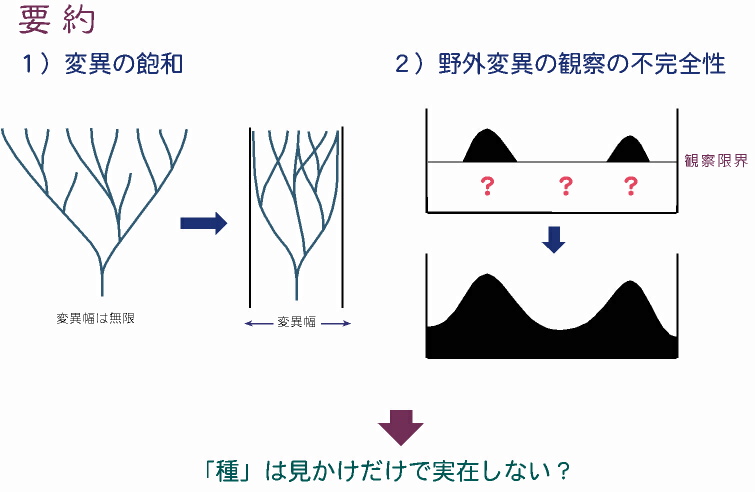 |
要 約 この講演の要旨は以下のとおりである。 1)原生生物の進化においては,分子・形態いずれにおいても,取り得る変異は無限ではなく「有限」である。そのため,系統進化がある程度続くと,途中で「変異の飽和」が起きてしまい,その形質(分子・形態)の違いと系統が相関しなくなる可能性が高い。いいかえると,各変異グループは単系統ではなく,多系統になるといえる。 2)多細胞生物は観察が比較的容易なため,野外変異をほぼ完全に把握できる。そこで,種の境界に「中間型」がいないことが確認できれば種が実在するといえる。しかし,原生生物の場合は,野外変異を完全に把握することはほぼ不可能である。この「観察限界」があることから,実際には中間型が存在し「種は実在しない」にもかかわらず,見かけ上,種とみなされている可能性がある。 |
 |
ゾウリムシ属の各形態種には接合型特異性の異なる複数のグループがある。 原生生物ゾウリムシ(Paramecium caudatum)では,従来,接合型の特異性が異なるグループは互いに生殖的に隔離されている(はず)なので,それぞれは別種である,と考えられてきた。しかし,私は接合型の遺伝解析を目的として接合型グループ間の雑種を作成したところ,その雑種に妊性がある(子孫を残す能力がある)ことを発見した(Tsukii & Hiwatashi 1983; Tsukii 1988)。これにより,グループ間では生殖的隔離が起きていない可能性が示唆された。 |
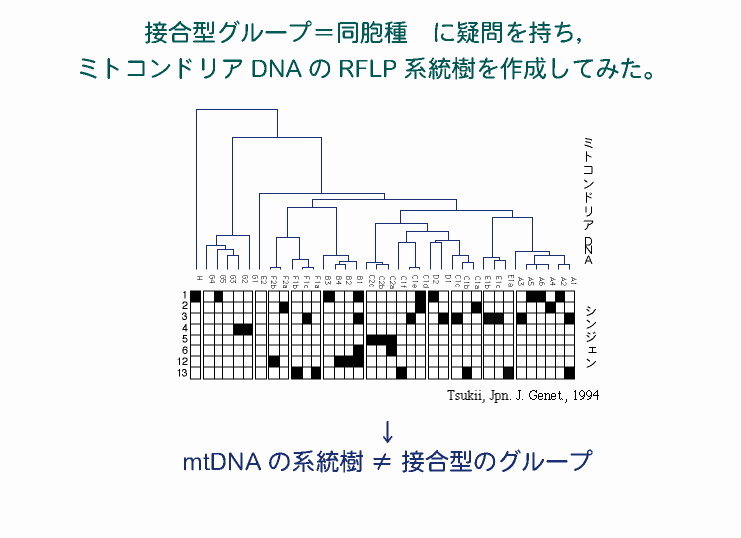 |
ミトコンドリアDNAのRFLP系統樹 その後,ゾウリムシから抽出したミトコンドリアDNA,および核DNAを使った分子系統樹を作成したところ,それらと接合型の分布はまったく一致しなかった(Tsukii 1994; Tsukii 1996)。 |
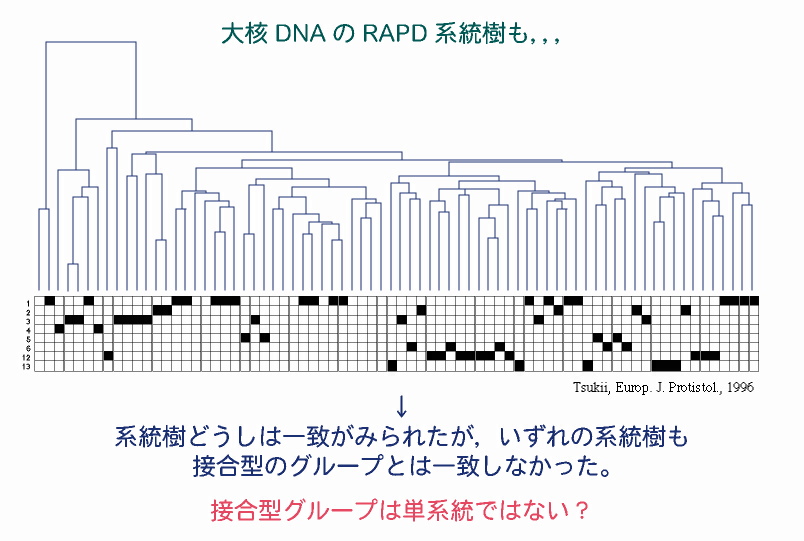 |
大核DNAのRAPD系統樹 同上 これらの結果は,各接合型グループは,いわゆる単系統(単一の祖先のみ由来する生物集団)ではなく多系統(系統的に異なる複数の祖先からなる集団)であることを強く示唆している。すなわち,ゾウリムシでは,祖先が異なるにもかかわらず,見かけ上,同じ接合型特異性を持つものが多数存在する(すなわち接合型に関して収斂進化が起きている)可能性が高い。 |
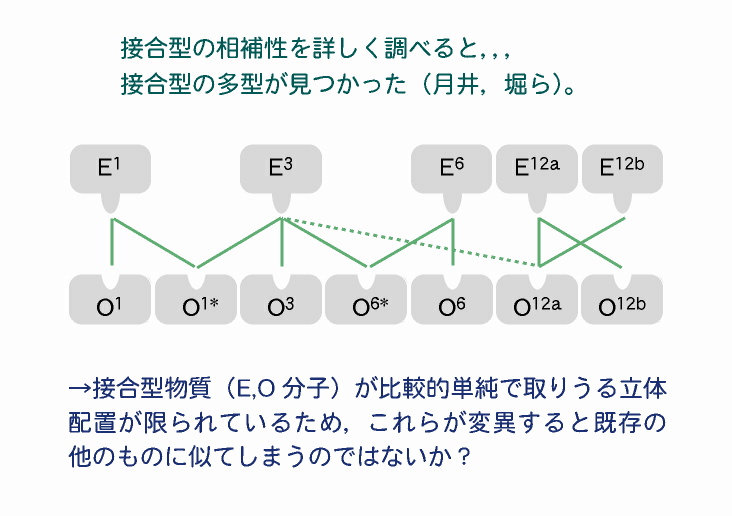 |
接合型の多型 実際すでに,野外からそれと思われる接合型の変異もいくつか発見されている(Tsukii 1988; 堀ら 1988)。 |
 |
少なくとも2億4000万年前からゾウリムシはいた! 近年の化石研究(Schönborn et al. 1999)から少なくとも2億4000万年前にはすでに今と形態的には変わらないゾウリムシが生息していたことが知られている。 |
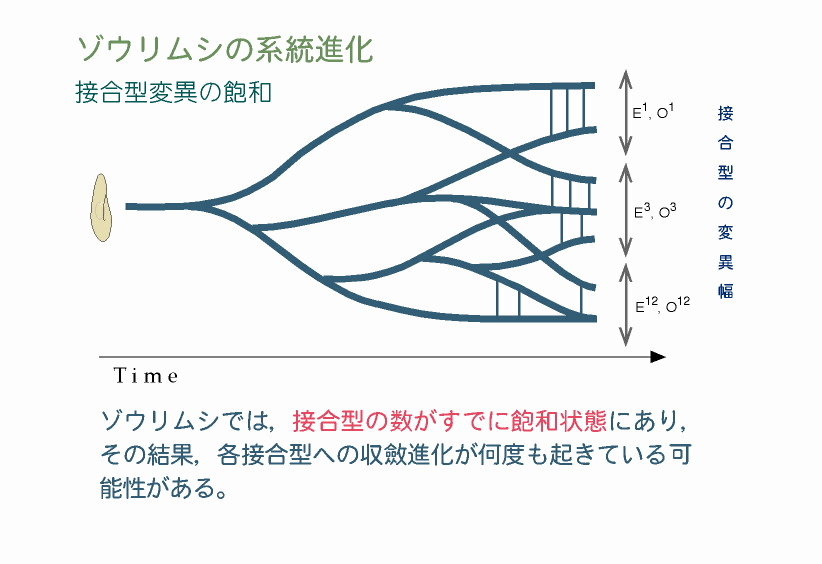 |
ゾウリムシの接合型変異はすでに飽和している!? P. caudatumでは形態的には単一種だが,既述した接合型の違いにより16の接合型グループに分かれていることが知られている。しかし,化石研究の結果で示された進化的起源の古さを考慮すると,突然変異により分化した接合型グループの数がわずか16というのは数が少なすぎる。この原因としては,接合型に関与する分子(タンパク質)の構造が比較的単純で,細胞接着に関与する接合型活性部位の取りうる立体配置が16種類しか存在しえなかったことが考えられる。そのため,16のグループが誕生した後に起きた接合型の変異は,すべて既存のいずれかの接合型に収斂してしまい16以上には数が増えなかったのであろう。これが,現在の接合型グループの多くが多系統になった理由と考えられる。 |
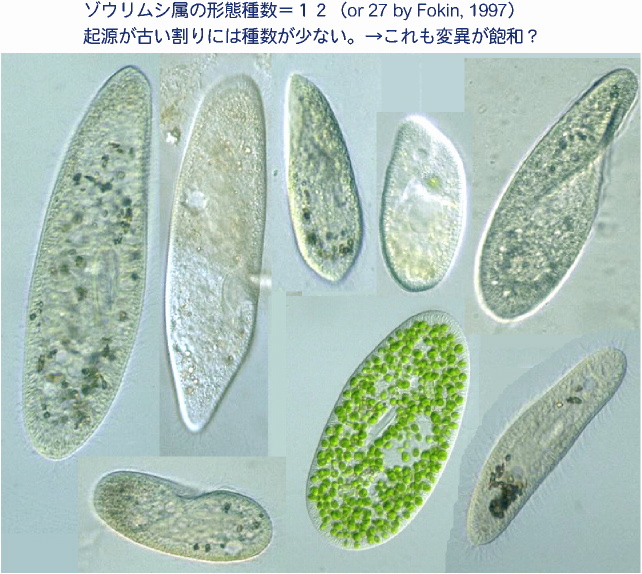 |
ゾウリムシは形態種が少なすぎる。これ以上変化できない!? 同様なことは,上位のカテゴリィであるゾウリムシ属(Paramecium)にもいえるのではないだろうか。現在,ゾウリムシ属に含まれる形態種(形態的特徴で区別された種)はわずか十数種しか知られていない。この数は,2億4000万年前からいる生物群としてはあまりに少ない。この場合は,ゾウリムシの細胞形態があまりに単純なため,取りうる「変異幅」が限られていて,形態レベルでは十数種以上には多様化できなかった可能性が考えられる。 |
 |
種内・属内の変異に興味を持ち他の原生生物も調べてみた。 以上のように,ゾウリムシでは,形態レベル,分子レベル(接合型の違い)のいずれにおいても,とりうる変異の幅が限られているため,その進化的起源の古さも手伝って,かなり以前から「変異の飽和」が起きていた可能性が高い。同様なことは,同じように形態が単純(識別可能な形態・機能的特徴が少ない)で,かつ,進化的起源が古い他の数多くの原生生物に当てはまるのではないかと考えられる。 |
 |
Mayorella属の核DNAを鋳型としたRAPD系統樹 そこで,既述したように,これまでデータベース制作の過程で得た様々な原生生物の種ごとの変異とその近縁種間の変異を調査するとともに,いくつかの属(Mayorella, Chilomonas, Frontonia)において属内の系統調査(具体的にはRAPD法による分子系統樹の作成)を行なった。その結果,形態的には多様性がみられないグループでも分子系統的には著しく多様性があるもの(Chilomonas)など,ゾウリムシの場合と同様に,他の原生生物においても形態レベルでの「変異の飽和」が起きていることを示唆する結果が得られた。 これは肉質虫類マヨレラ属の核DNAを使ったRAPD系統樹である。マヨレラ属は形態が多様で,種を識別するのが大変にむずかしい。一応,形態が似たもの(よって同一種と判断されたもの)は系統的に近い場合(M. leidyi)もあったが,マヨレラよりも他の属(Korotnevella)に近いものなど全体としては形態的類似性と分子系統はあまり一致しなかった。 |
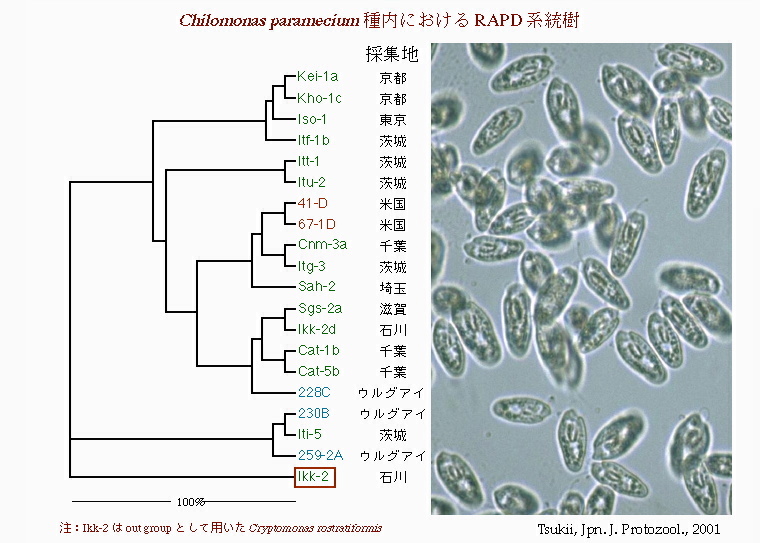 |
Chilomonas paramecium種内におけるRAPD系統樹 これは鞭毛虫類キロモナス(Chilomonas paramecium)の核DNAを使った種内のRAPD系統樹である。C. parameciumは,土壌や池沼の泥水等に広く生息する種類で,採集の際,もっとも頻繁に発見される。にもかかわらず,形態的な変異はほとんどみられず,日本で発見されているキロモナス属の種類はこのC. paramecium,わずか一種類のみである。 しかし,分子系統樹を作成すると種内の変異量は,最大100%に達し,前のマヨレラ属以上であった。このことは,C. parameciumは進化的起源が相当古い種類であるが,なんらかの理由で形態的な多様化が抑制されてきたことを強く示唆している。 |
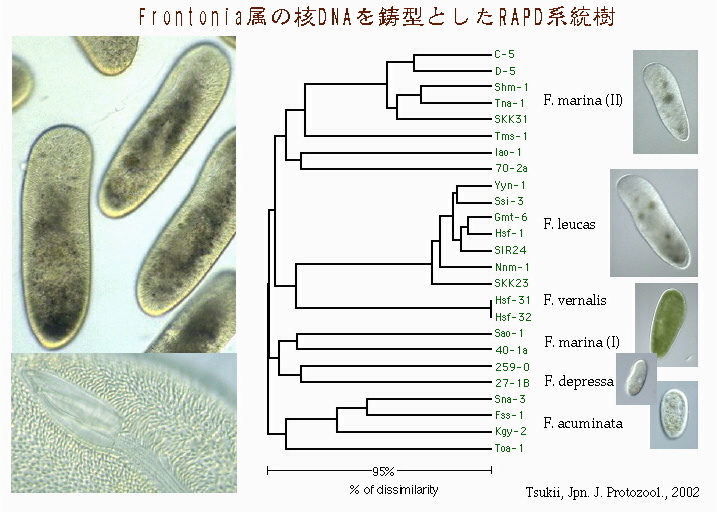 |
Frontonia属の核DNAを鋳型としたRAPD系統樹 これは繊毛虫類フロントニア(Frontonia)属の核DNAを使ったRAPD系統樹である。意外だったのは,野外にもっとも多く生息する大型種,F. leucasは,採集地が離れたものでもほとんど分子レベルでは違いがみられなかったことである。しかし,属全体でみると,やはり形態的類似性と分子系統との間に相関はみられなかった。とくに中型種 F. marinaは多様で,他の種と系統的に入り交じっていた。これらのことから,F. leucasは比較的近年になって進化し,急速に分布を拡大した種であることと,一方,F. marinaとされるグループは,分子系統的には「多系統」であるといえる。 |
 |
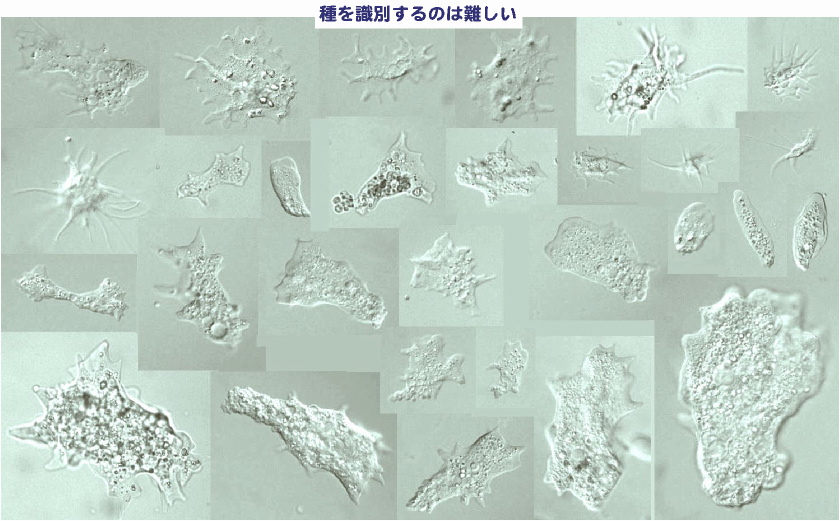
種を識別するのはむずかしい。(Saccamoeba の場合) 考えてみれば,単細胞の原生生物は,多細胞の我々よりも進化的起源の古いものがたくさんいても不思議はない(既述したように化石の証拠もある)。しかし,原生生物の多くは小さく形態が単純なものが多い。ミカヅキモなどの微小藻類の仲間は微細な形態的特徴を識別しやすいので比較的多数の(形態)種が報告されているが,いわゆる「原生動物」と呼ばれてきたものの多くは,絶えず形態変化していることもあって細かく種を区別することができず,その結果,報告されている種の数も少ない傾向にある。 ここに示したのは,アメーバの仲間(裸性根足虫類)であるサッカメーバ(Saccamoeba)属の画像だが,野外採集をすると無限ともいえるほど様々な大きさ,形のものが見つかってくる。そのため,これらを種に分けるのは至難の技といえる。 |
 |
種を識別するのはむずかしい。(Mayorella の場合) Saccamoeba属以上に頻繁に見つかるのが,このマヨレラ属である。池沼・田圃・下水から採集したサンプルには必ずといってよいほど発見される。あまりに多いので撮影したのがごくごく一部にすぎないが,それでもこのように様々な形態のものが見つかる。サッカメーバ同様,これらを「種に分ける」というのはほとんど絶望的ともいえよう。 |
 |
野外変異観察の不完全性 以上のように,原生生物では,識別可能な細胞形態や機能(接合型等)に基づいて区別された「種」は,分子系統学的には単系統ではなく,したがって実在する種とは呼べないことがわかってきた。それでは原生生物では何を根拠に種が存在(実在)するといえるのだろうか? 一般に,教科書的には,種とは「互いに生殖的に隔離された生物集団」であると定義(これを生物学的種という)され,この機能的な定義により種には「実在性」があると考えられている。しかし,自然界には有性生殖を行なわない生物も多数存在するので,上記の生物学的種概念には普遍性がなく,種の遍在性を主張する根拠にはなりえない。 また,有性生殖を行なう生物であっても,実際に生殖的隔離の有無で種を区別することは希である。通常,種の存在を確認する重要な手がかりとなるのは,種間に存在する「変異の不連続性」である。すなわち,中間的な形質(形態と性質)をもつ個体がいないことが種を区別する上での実質的な判断基準となっているのである。この「変異の不連続性」は有性生殖をする・しないに関わらず確認できるので,種の遍在性の重要な証拠とみなされている。 たしかに多細胞生物(動物&植物&菌類)では,個体の観察が比較的容易なため,各種ごとの変異の全体像がおおよそ把握できている。これらの「観察事実」に基づいて,変異の不連続性が確認され,種が実在するとみなされている。しかし,原生生物や他の微生物では,研究の出発点となる野外変異の観察そのものが非常に困難であり,相当な労力を費やしても,我々は野外変異のごく一部しか捉えることができない。個体数の多い変異は見つかりやすいが,個体数の比較的少ないものは,通常の観察ではほとんど,あるいはまったく発見できない。 |
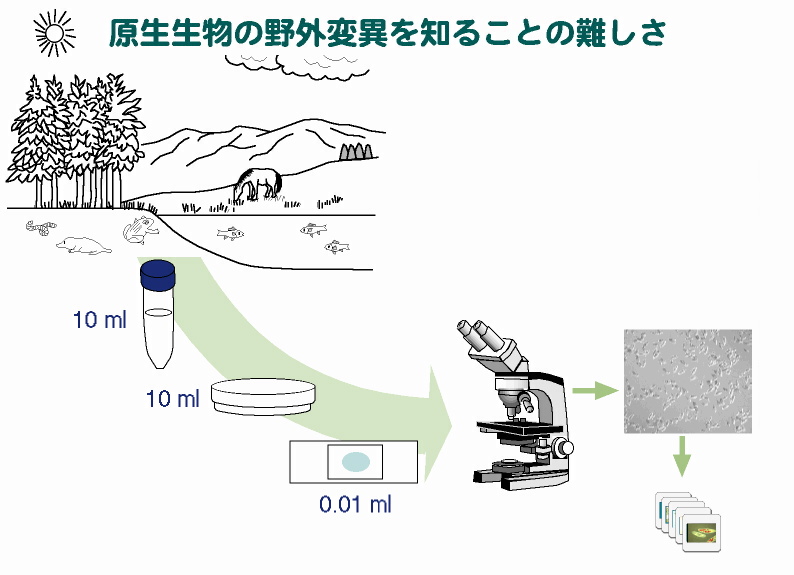 |
原生生物の野外変異を知ることの難しさ 顕微鏡で一度に観察できるサンプルの量は多くても0.1 mlかそれ以下である。したがって,例えば 1 tの水の中に1000匹の原生生物がいたとして,その中のわずか1匹を発見するだけでも単純計算で最大2000時間も費やさなければならない。眼の前に1000匹(個体)も生物がいれば,動物や植物なら絶対に見逃すことはないが,原生生物の場合はほとんどいないのと同然なのである。 また,同種の細胞が1サンプルに何万匹いたとしても,その大部分は1〜数個の細胞が分裂してできたクローンである場合がほとんどである。そのため,別の「個体」を観察するには,採集地点を変えて採集をくり返さなければならない。すなわち,多細胞生物ならば,(種類にもよるだろうが)一度の採集(ないし野外観察)で100個体程度を調べるのはさほど難しくはないが,原生生物の場合は,100個体を観察するには,100箇所以上の異なる地域からサンプルを採集しなければならないのである。 |
 |
過去五年間の観察で一度しか遭遇しなかった生物たち ここに示したのは,私が過去およそ5年間,かなり頻繁に野外採集を行ない,写真撮影(現在までに6400サンプルを撮影した)を行なってきたにもかかわらず,その間に,たった一度しかお目にかからなかった生物達である。 5年間に必死に探しても一度しか観察できないとなると,多細胞生物ならば絶滅危惧種に指定されてしまいそうだが,原生生物の場合は,前に示した理由で,そうではないのである(1 tの水の中に1000匹いても,実際には,ほとんど観察できない)。むしろ,5年の間に一度観察できたということは,その生物はかなり個体数の多い種類に属するのかも知れない。 |
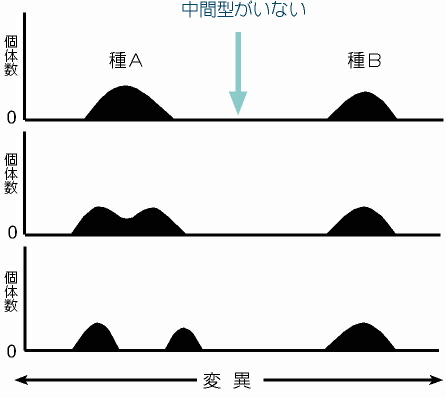 |
観察限界を考慮した進化パターン図1 この図は従来の進化のイメージである。すなわち,各種は様々な変異をもった個体の集合ではあるが,他の種との間には中間型がいないことで明瞭に区別される。1つの山(種)が隔離などの影響で変異幅の異なる2つの山に分かれることで種の分化が起こる。 |
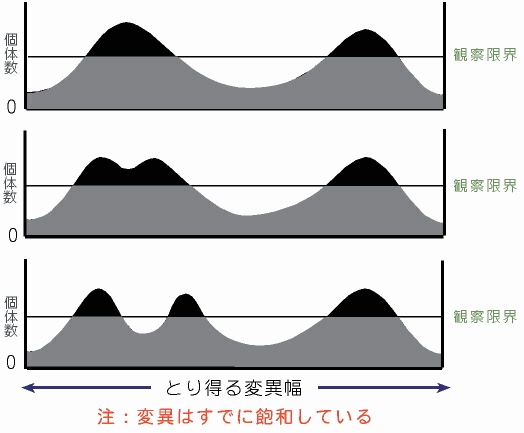 |
観察限界を考慮した進化パターン図2 一方,観察限界がある場合は,かりに中間型が存在して変異が連続していたとしても,見かけ上は前の図と同じになる。しかし,1つの山が2つに分かれたとしても,それは単にこれまで観察できた変異の個体数が減って観察できなくなり,替わりに従来は観察できなかった変異の数が増えて観察可能になっただけ,かも知れない。 このような「観察限界」の存在と,既述した「変異の飽和」を考慮すると,実際には起きていない種進化があたかも起きているかのごとく見えてしまったり,さらにはこれまで存在していなかった種が突然現れたり,その逆も起こり得るはずである。 |
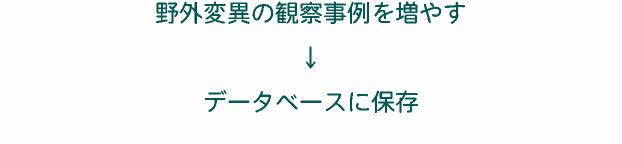 |
我々はもっと野外観察をすべきだ 微生物の種は,通常,野外から採集してきた細胞を実験室内で培養して,それを詳しく調べて種の記載をすることが多い。多少の野外変異も調べられるが,それは多細胞生物に比べれば圧倒的に少なく,網羅的ではない。そのため,文献によって各種の変異幅(細胞長など)はまちまちである。中には,以前の文献にある記録に新規データを追加して変異幅を記載している文献もあるが,その場合は時代とともに変異幅が大きくなる傾向がみられる。 原生生物の種について考えるためには,我々はもっと数多くの野外変異を知る必要がある。しかし,問題は,原生生物のほとんどは「保存標本」が作れないという点にある。従来,原生生物の野外観察が十分に行なわれなかったのは,このためもあっただろう。そこで,その問題を解決し,野外観察を行なうために有効なのが,私たちが現在行なっている画像データベース(我々はこれをデジタル標本箱と呼んでいる)なのである。 |
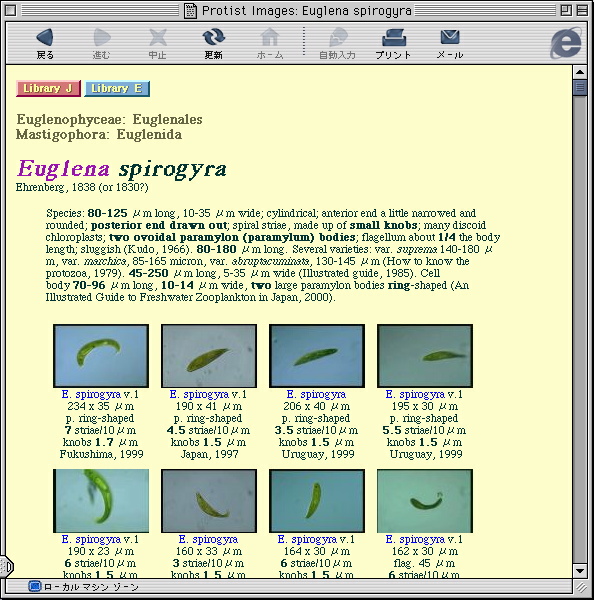 |
画像データベースの一画面 これは原生生物情報サーバの「デジタル標本館」にあるEuglena 属の index pageである。各種名をクリックすると,その種のweb pageへ移動する。そこには多数の標本web pagesへのリンクがあり,各標本web pageにはこれまで野外から採集されたサンプルの画像がある。 |
 |
Euglena spirogyra の種内変異(1) これはその中の一つ。Euglena spirogyraの野外変異。ここにあるのは文献にある変異幅内に収まるサンプルの画像だが,,。 |
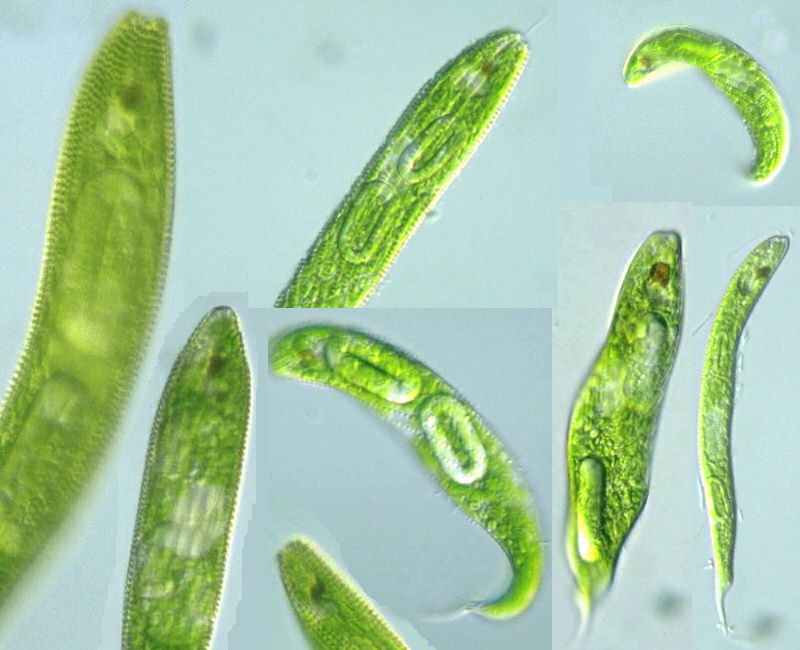 |
Euglena spirogyra の種内変異(2) 採集サンプルが増えるにつれ,文献にある種の特徴の変異幅を越えたものも見つかってきた。今後,さらに採集を続けていけばどこまで変異の幅が広がるかが問題。 |
 |
Euglena mutabilis の種内変異(1) 同様なことは,Euglena mutabilisでも起きた。文献的にはこの程度の変異までが紹介されているが,さらに調べると,,。 |
 |
Euglena mutabilis の種内変異(2) さらに大きなものや,非常に小さなものまで見つかってきた。小さなものは決して痩せ細っている訳ではない。その証拠に,これらは無菌培養しているもので,栄養が十分すぎるくらいある条件で撮影したものであり,かれらはこれ以上には決して大きくならない(右下図)。 |
 |
考えられる原生生物の進化イメージ 原生生物では種の境界が存在しない可能性がある。とすれば進化はこのような図で表現せざるをえないはずである。図の濃淡は各変異の個体数を表している。ただし,観察限界を考慮して観察できない部分を白抜きにした場合は,従来の「系統樹」に変化する。 また,途中から変異の飽和が起こるために,どの変異領域をグループ化してもそれは単系統ではなく多系統である可能性が高い。 |
 |
まとめ 原生生物は,識別可能な形態的・機能的特徴が少ないため,とり得る変異幅(種類)が限られている。多くは進化的起源が古いので,すべての変異が出尽くしている(変異が飽和している)可能性が高い。 原生生物は多数の野外変異を観察するのが難しい(観察限界がある)。このため,種の境界の有無が不明だが,観察量を増やした場合,種の境界が存在しなくなる可能性がある。 その場合,我々が「種」と呼んでいるものは見かけだけで,種は実在しないことになる。 |