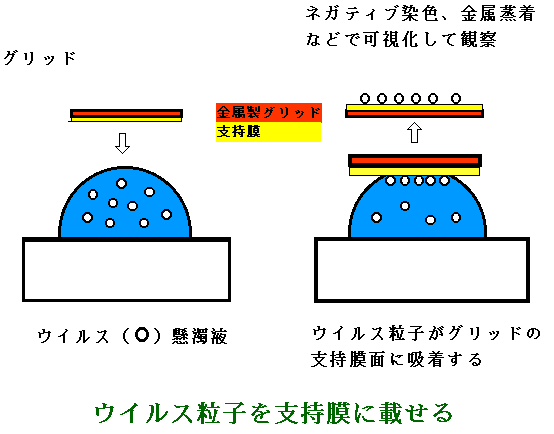
ウイルス等の入った液を少量(20~100μl)とってガラス等の上に液滴とし、この上に支持膜の張ってある面を下にしてグリッドを浮かせると、液滴中のウイルス粒子が拡散しながら支持膜に吸着する。(図2)これをネガティブ染色や金属蒸着をしてTEMで観察する。

|
光学顕微鏡の場合に、試料を載せるために使われるスライドグラスに当たるものは、スライドグラスのようなガラスが電子線を透過させないので、透過型電子顕微鏡(TEM)では、グリッドと呼ばれるものを使う。グリッドは直径3mmの金属板で、図1のように小さな孔が沢山あいている。この上にコロジオン、パロジオン、ホルムバール等のプラスチックの薄膜を張り、さらに炭素粒で補強する(カーボン蒸着)。カーボン蒸着は真空蒸着機中で炭素棒を熱して炭素粒を飛散させて行う。電子線は、薄いプラスチックは透過するので、ここに試料を載せて観察する。 ウイルス等の入った液を少量(20~100μl)とってガラス等の上に液滴とし、この上に支持膜の張ってある面を下にしてグリッドを浮かせると、液滴中のウイルス粒子が拡散しながら支持膜に吸着する。(図2)これをネガティブ染色や金属蒸着をしてTEMで観察する。 |
図1
|
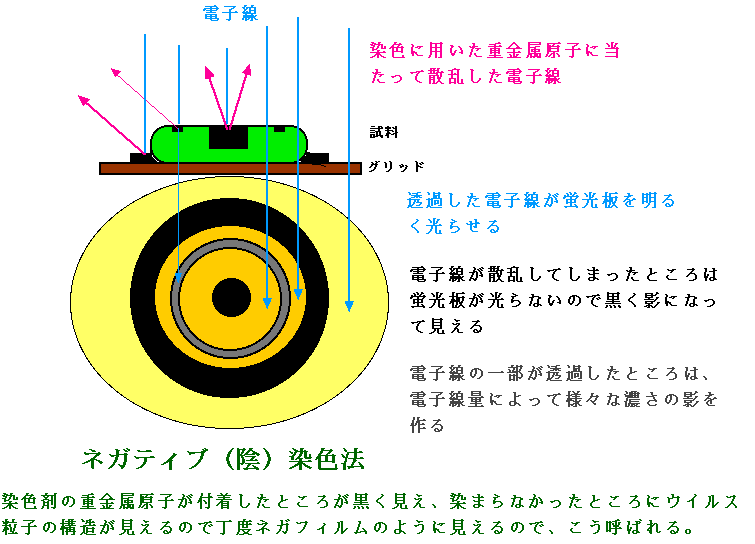
| ウイルスを含めて生物の体は主に、水素(H)、炭素(C)、窒素(N)、酸素(O)等の軽元素で出来ている。これらの元素は小さいので電子を散乱させにくい。試料に電子が当たっても散乱されずに透過してしまうばかりではコントラストがつかないので、元素が大きな鉛やウランなどの重金属で染色して電子を散乱させるようにしてコントラストのはっきりした像を得る。ウイルス粒子の観察には、ネガティブ染色法が良く用いられる。(図3)グリッドに載っている試料に重金属溶液を加え濾紙で余分な染色液を吸い取ると、重金属原子が支持膜や支持膜と試料の間、試料の凹凸の凹部などに残留する。その結果、重金属が残った部分は電子が散乱され影となり、試料の凸部は電子が透過するので影の縁取りの中に明るく浮き上がって見える。試料そのものが染色されるのではなく、試料凹部や試料のない部分に重金属が残ってネガフィルムのような逆のコントラストに染色されているように見えるのでネガティブ染色という。ネガティブ染色には、モリブデン(Mo)、タングステン(W)、ウラン(U)等の重金属が用いられる。ウイルス粒子の殻の構成蛋白質までもが明瞭に観察できることがある。固定、包埋などの前処理を必要とせず簡単に観察できる割には得られる情報量の大きな便利な方法である。感染性のある危険なウイルスでは、感染性を失わせる不活化処理(紫外線照射、グルタールアルデヒド固定など)を行ってから染色する。なお、図4はネガティブ染色法によって観察されたアデノウイルスの写真である。 |
図4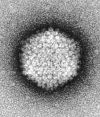
|
| ウイルス粒子や蛋白質、核酸(遺伝子など)の観察には、金属蒸着法も用いられる。真空中でタングステン線に電圧をかけるとタングステン線が白熱する。(白色電球と同じ事)このとき、タングステン線に白金ーパラジウム合金(白金Pt80 ; パラジウムPd20)の細い線(太さ0.1mm)を巻いておくと、Pt-Pd合金が細かい粒子となって飛散する。この粒子が、ウイルス粒子表面や核酸に付着すると、よく電子線を散乱し表面の凹凸が強調されて観察されるようになる。飛散する粒子が細かいほど、微細な表面構造が可視化される。真空蒸着機を使って行う。Pt-Pd合金の他、白金、金、タングステンなどを蒸着する事もある。なお、図5は金属蒸着法によって観察されたカイコ細胞質多角体病ウイルスの写真である。 |
図5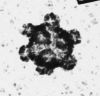
|